Чему равен дыхательный коэффициент при окислении углеводов. Дыхательный коэффициент. Дыхательный коэффициент при работе
Отношение объема выделенной двуокиси углерода к объему поглощенного кислорода называется дыхательным коэффициентом.
ДК = СО 2 (л)/О 2 (л)
Дыхательный коэффициент характеризует тип питательных веществ, преимущественно окисляемых в организме на момент его определения. Его рассчитывают, исходя из формул химических окислительных реакций.
Для углеводов :
С 6 Н 12 О 2 + 6О 2 о - 6СО 2 + 6Н 2 О;
ДК = (6 объемов СО 2)/(6 объемов О 2) = 1
Для жиров :
2С 15 Н48,О 6 + 145О 2 о - 102СО 2 + 98Н 2 О;
ДК = (102 объема СО 2)/(145 объемов О 2) = 0,703
Для белков расчет представляет определенную трудность, так как белки в организме окисляются не полностью. Некоторое количество азота в составе мочевины (NH 2) 2 CO 2 выводится из организма с мочой, потом и фекалиями. Поэтому для расчета ДК при окислении белка следует знать количество белка, поступившего с пищей, и количество экскретированных азотсодержащих «шлаков». Установлено, что для окисления углерода и водорода при катаболизме белка и образования 77,5 объема двуокиси углерода необходимо 96,7 объема кислорода. Следовательно, для белков:
ДК = (77,5 объема СО 2)/(96,7 объема О 2) = 0,80
При смешанной пище дыхательный коэффициент составляет 0,8-0,9.
Дыхательный коэффициент при мышечной работе. Главным источником энергии при интенсивной мышечной работе являются углеводы. Поэтому во время работы ДК приближается к единице.
Сразу по окончании работы ДК может резко повыситься. Это явление отражает компенсаторные процессы, направленные на удаление из организма избытка двуокиси углерода, источником которого являются так называемые нелетучие кислоты.
Через некоторое время по завершении работы ДК может резко снизиться по сравнению с нормой. Это связано с уменьшением выделения двуокиси углерода легкими вследствие компенсаторной задержки его буферными системами крови, предотвращающими сдвиг рН в основную сторону.
Примерно через час после завершения работы ДК становится нормальным.
Калорический эквивалент кислорода. Определенному дыхательному коэффициенту соответствует определенный калорический эквивалент кислорода, т.е. количество тепла, которое освобождается при полном окислении 1г питательного вещества (до конечных продуктов) в присутствии 1л кислорода.
Калорический эквивалент кислорода при окислении белков равен 4,8 ккал (20,1 кДж), жиров - 4,7 ккал (19,619 кДж), углеводов - 5,05 ккал (21,2 кДж).
Первоначально газообмен у человека и животных определяли методом Крога в специальных камерах закрытого типа (респираторная камера М.Н. Шатерникова).
В настоящее время полный газовый анализ проводят открытым респираторным методом Дугласа-Xолдейна. Метод основан на сборе выдыхаемого воздуха в специальный приемник (воздухонепроницаемый мешок) с последующим определением общего его количества и содержания в нем кислорода и двуокиси углерода при помощи газоанализаторов.
№ 51 Основной обмен и методы его определения. Условия определения основного обмена и факторы, влияющие на его величину. Специфическое динамическое действие пищи. Закон поверхности М. Рубнера.
Основной обмен - минимальное количество энергии, необходимое для обеспечения нормальной жизнедеятельности в условиях относительного физического и психического покоя. Эта энергия расходуется на процессы клеточного метаболизма, кровообращение, дыхание, выделение, поддержание температуры тела, функционирование жизненно важных нервных центров мозга, постоянную секрецию эндокринных желез.
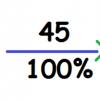
Печень потребляет 27 % энергии основного обмена, мозг - 19 %, мышцы - 18 %, почки - 10 %, сердце - 7 %, все остальные органы и ткани - 19 %.
Методы определения основного обмена.
Расчет основного обмена по таблицам . Специальные таблицы дают возможность по росту, возрасту и массе тела определить средний уровень основного обмена человека. При сопоставлении этих величин с результатами, полученными при исследовании рабочего обмена с помощью приборов, можно вычислить разницу, эквивалентную затратам энергии для выполнения работы.
Вычисление основного обмена по гемодинамическим показателям (формула Рида). Расчет основан на взаимосвязи между артериальным давлением, частотой пульса и теплопродукцией организма. Формула дает возможность вычислить процент отклонения величины основного обмена от нормы. Допустимым считается отклонение ±10 %.
ПО = 0,75 (ЧСС + ПД 0,74) - 72,
где ПО - процент отклонений; ЧСС - частота сердечных сокращений
(пульс); ПД - пульсовое давление.
Для определения соответствия основного обмена нормативным данным по гемодинамическим показателям существуют специальные номограммы.
Расход энергии в состоянии покоя различными тканями организма неодинаков. Более активно расходуют энергию внутренние органы, менее активно - мышечная ткань. Интенсивность основного обмена в жировой ткани в 3 раза ниже, чем в остальной клеточной массе организма. Люди с низкой массой тела производят больше тепла на 1 кг массы тела, чем с высокой. Если рассчитать энегoвыделение на 1 м 2 поверхности тела, то эта разница почти исчезает. Согласно еще одному правилу Рубнера, основной обмен приблизительно пропорционален поверхности тела для разных видов животных и человека.
Отмечены сезонные колебания величины основного обмена - повышение его весной и снижение зимой. На величину основного обмена влияют предшествующая мышечная работа, состояние желез внутренней секреции.
Условия определения основного обмена.
Любая работа - физическая или умственная, а также прием пищи, колебания температуры окружающей среды и другие внешние и внутренние факторы, изменяющие уровень обменных процессов, влекут за собой увеличение энерготрат.
Поэтому основной обмен определяют в строго контролируемых, искусственно создаваемых условиях: утром, натощак (через 12-14 ч после последнего приема пищи), в положении лежа на спине, при полном расслаблении мышц, в состоянии спокойного бодрствования, в условиях температурного комфорта (18-20 °С). За 3 сут до исследования из рациона исключают белковую пищу. Выражается основной обмен количеством энергозатрат из расчета 1 ккал на 1 кг массы тела в час .
Факторы, определяющие величину основного обмена. Основной обмен зависит от возраста, роста, массы тела, пола человека. Самый интенсивный основной обмен в расчете на 1 кг массы тела отмечается у детей (у новорожденных - 53 ккал/кг в сутки, у детей первого года жизни - 42 ккал/кг). Средние величины основного обмена у взрослых здоровых мужчин составляют 1300-1600 ккал/сут; у женщин эти величины на 10 % ниже. Это связано с тем, что у женщин меньше масса и поверхность тела.
Специфическое динамическое действие пищи - повышение энерготрат организма, обусловленное приемом, перевариванием и усвоением пищи. Специфическое динамическое действие пищи состоит в том, что на переваривание пищи, даже в отсутствии мышечной активности, также расходуется энергия. При этом наибольший расход вызывает переваривание белков. Белки обладают максимально усиливающим действием на обмен веществ, они увеличивают его на 40%, углеводы и жиры увеличивают его всего на 5%. При обычном питании суточный расход на специфическое динамическое действие пищи у взрослого человека составляет около 200 калорий.
Закон поверхности тела Рубнера. Зависимость интенсивности основного обмена от площади поверхности тела была показана немецким физиологом Рубнером для различных животных. Согласно этому правилу, интенсивность основного обмена тесно связана с размерами поверхности тела: у теплокровных организмов, имеющих разные размеры тела, с 1 м 2 поверхности рассеивается одинаковое количество тепла.
Таким образом, закон поверхности тела гласит: энергетические затраты теплокровного организма пропорциональны площади поверхности тела.
С возрастом величина основного обмена неуклонно снижается. Средняя величина основного обмена у здорового человека равна приблизительно 1 ккал/(кг-ч).
№ 52 Рабочий обмен энергии. Энергетические затраты организма при различных видах труда. Методы определения рабочего обмена.
Общий расход энергии человеком зависит от состояния организма и мышечной деятельности.
Мышечная работа сопряжена со значительными затратами энергии (рабочий обмен энергии ), с одной стороны, и увеличением теплопродукции - с другой. У спокойно лежащего человека теплопродукция составляет 35 ккал/(гм 2). Если исследуемый принимает сидячее положение,- на 42 %; в положении стоя - на 70 %, а при спокойной неторопливой ходьбе теплопродукция увеличивается на 180 %. При мышечных нагрузках средней интенсивности КПД работы мышц составляет около 24 %. Из всего количества энергии, расходуемой работающими мышцами, 43 % затрачивается на активацию сокращения, и вся эта энергия переходит в тепло. Только 57 % из общего количества энергии идет на рабочее сокращение.
Разность между энергозатратами при физической нагрузке и энергозатратами основного обмена составляет рабочую прибавку, которая тем больше, чем интенсивнее работа. Рабочая прибавка - это вся остальная энергия, которую тратит организм в течение суток на физическую и умственную активность.
Сумма основного обмена и рабочей прибавки составляет валовый обмен. Сумма валового обмена и специфического динамического действия пищи называется общим обменом.Предельно допустимая по тяжести работа для данного человека, постоянно выполняемая им в течение длительного времени, не должна превышать по энергозатратам уровень основного обмена более чем в 3 раза. При кратковременных нагрузках энергия выделяется за счет окисления углеводов.
При длительных мышечных нагрузках в организме расщепляются преимущественно жиры, обеспечивая 80 % потребной энергии. У тренированных спортсменов энергия мышечных сокращений обеспечивается исключительно за счет окисления жиров. У человека, занимающегося физическим трудом, энергетические затраты возрастают пропорционально интенсивности труда.
По энергетическим затратам все профессии разделены на несколько групп, каждая из которых характеризуется своим суточным расходом энергии.
Коэффициент физической активности. Объективным физическим критерием, определяющим адекватное количество расходования энергии для конкретных профессиональных групп людей, является коэффициент физической активности (отношение общих энерготрат на все виды жизнедеятельности к величине основного обмена, т.е. расходу энергии в состоянии покоя). Величины коэффициента физической активности одинаковы для мужчин и женщин, но в связи с меньшей величиной массы тела у женщин и соответственно основного обмена энерготраты мужчин и женщин в группах с одним и тем же коэффициентом физической активности различны.
Группа I - работники преимущественно умственного труда: научные работники, студенты гуманитарных специальностей. Очень легкая физическая активность; коэффициент физической активности 1,4; расход энергии 1800-2450 ккал/сут.
Группа II - работники, занятые легким физическим трудом: водители трамваев, троллейбусов, работники сферы обслуживания, медицинские сестры, санитарки. Легкая физическая активность; коэффициент физической активности 1,6; расход энергии 2100- 2800 ккал/сут.
Группа III - работники средней тяжести труда: слесари, настройщики, водители автобусов, врачи-хирурги. Средняя физическая активность; коэффициент физической активности 1,9; расход энергии 2500-3300 ккал/сут.
Группа IV - работники тяжелого физического труда: строительные рабочие, металлурги. Высокая физическая активность; коэффициент физической активности 2,2; расход энергии 2850- 3850 ккал/сут.
Группа V - работники особо тяжелого труда, только мужчины: механизаторы, сельскохозяйственные рабочие в посевной и уборочный периоды, горнорабочие, вальщики леса, бетонщики, каменщики, землекопы, грузчики немеханизированного труда, оленеводы и др. Очень высокая физическая активность; коэффициент физической активности 2,5; расход энергии 3750-4200 ккал/сут.
Для каждой группы труда определены средние величины сбалансированной потребности здорового человека в энергии и пищевых веществах, которые несколько различаются для мужчин и женщин.
№ 53 Температура тела человека и ее суточные колебания. Тепловой баланс гомойотермного организма. Температурная схема тела человека. Методы измерения температуры тела человека.
Гомойотермия. В процессе эволюции у высших животных и человека выработались механизмы, способные поддерживать температуру тела на постоянном уровне независимо от температуры окружающей среды. Температура внутренних органов у них колеблется в пределах 36-38 °С, способствуя оптимальному течению метаболических процессов, катализируя большинство ферментативных реакций и влияя в определенных границах на их скорость.
Постоянная температура необходима и для поддержания нормальных физико-химических показателей - вязкости крови, ее поверхностного натяжения, коллоидно-осмотического давления и др. Температура влияет и на процессы возбуждения, скорость и интенсивность сокращения мышц, процессы секреции, всасывания и защитные реакции клеток и тканей.
Гомойотермные организмы выработали регуляторные механизмы, делающие их менее зависимыми от окружающих условий. Они способны избегать перегревания при слишком высокой и переохлаждения при слишком низкой температуре воздуха.
Оптимальная температура тела у человека составляет 37 °С; верхняя летальная температура - 43,4 °С. При более высокой температуре начинается внутриклеточная денатурация белка и необратимая гибель; нижняя летальная температура составляет 24 °С. В экстремальных условиях резких изменений окружающей температуры гомойотермные животные реагируют реакцией стресса (температурный - тепловой или холодовой - стресс). С помощью этих реакций такие животные поддерживают оптимальный уровень температуры тела. Гомойотермия у человека вырабатывается в течение жизни.
Температура тела человека, а также высших животных подвержена более или менее правильным суточным колебаниям даже при одних и тех же условиях питания и физической активности.
Температура тела днем выше, чем ночью, и в течение суток колеблется в пределах 0,5-3 °С, снижаясь до минимального уровня в 3-4 ч утра и достигая максимума к 16-18 ч вечера. Суточный ритм температурной кривой не связан непосредственно со сменой периодов активности и покоя, поскольку он сохраняется и в том случае, если человек постоянно находится в полном покое. Этот ритм поддерживается без каких-либо внешних регулирующих факторов; он присущ самому организму и представляет собой истинно эндогенный ритм.
У женщин выражены месячные циклы колебаний температуры тела. температура повышается после приема пиши (специфическое динамическое действие пищи), при мышечной работе, нервном напряжении.
Температурная схема тела , которая определяется различным уровнем обмена веществ в разных органах. Температура тела в подмышечной впадине - 36,8 °С, на ладонных поверхностях руки - 25-34 °С, в прямой кишке - 37,2-37,5 °С, в ротовой полости - 36,9 °С. Самая низкая температура отмечается в пальцах нижних конечностей, а самая высокая - в печени.
Вместе с тем даже в одном и том же органе существуют значительные температурные градиенты, а ее колебания составляют от 0,2 до 1,2 °С. Так, в печени температура равна 37,8-38 °С, а в мозге - 36,9-37,8 °С. Значительные температурные колебания наблюдаются при мышечной нагрузке. У человека интенсивная мышечная работа приводит к повышению температуры сокращающихся мышц - на 7 °С.
При купании человека в холодной воде температура стопы падает до 16 °С без каких-либо неприятных ощущений.
Индивидуальные особенности температурной схемы тела:
Здоровый человек имеет относительно постоянную температурную схему тела;
Особенности температурной схемы генетически детерминированы, в первую очередь индивидуальной интенсивностью метаболических процессов;
Индивидуальные особенности температурной схемы тела определяются влияниями гуморальных (гормональных) факторов и тонусом вегетативной нервной системы;
Температурная схема тела совершенствуется в процессе воспитания, определяется образом жизни и особенно закаливанием. Вместе с тем она динамична в известных пределах, зависит от особенностей профессии, экологических условий, характера и других факторов.
№ 54 Механизмы теплопродукции. Обмен веществ как источник образования тепла. Роль отдельных органов в теплопродукции и регуляция этого процесса.
Центры теплообразования. В области латерально-дорсального гипоталамуса обнаружены центры теплообразования. Их разрушение приводит к тому, что животные утрачивают способность поддерживать постоянство температуры тела в условиях пониженной температуры окружающей среды. Температура их тела в этих условиях начинает падать, и животные переходят в состояние гипотермии. Электрическое раздражение соответствующих центров гипоталамуса вызывает у животных следующий синдром: 1) сужение поверхностных сосудов кожи. Вазоконстрикция достигается активацией симпатических центров заднего гипоталамуса.; 2) пилоэрекцию- реакция выпрямления волос тела.; 3) мышечную дрожь - увеличивает величину теплопродукции в 4–5 пять раз. Двигательный центр дрожи располагается в дорсомедиальной части заднего гипоталамуса. Он тормозится повышенной внешней температурой и возбуждается при её понижении. Импульсы из центра дрожи вызывают генерализованное повышение мышечного тонуса. Повышенный мышечный тонус приводит к возникновению ритмических рефлексов с мышечных веретён, что и вызывает дрожь; 4) увеличение секреции надпочечников.
Взаимодействие центров терморегуляции. Между центрами теплоотдачи переднего гипоталамуса и центрами теплопродукции заднего гипоталамуса существуют реципрокные взаимоотношения. При усилении активности центров теплопродукции тормозится деятельность центров теплоотдачи и наоборот. При снижении температуры тела включается активность нейронов заднего гипоталамуса; при повышении температуры тела активируются нейроны переднего гипоталамуса.
Механизмы теплопродукции. При снижении температуры окружающей среды эфферентная импульсация от нейронов заднего отдела гипоталамуса распространяется на α-мотонейроны спинного мозга. Эти влияния приводят к сокращению скелетных мышц. При сокращении мышц возрастает гидролиз АТФ. Вследствие этого увеличивается произвольная мышечная активность.
Одновременно при охлаждении возрастает так называемый терморегуляционный тонус мышц. Терморегуляционный тонус представляет своеобразную микровибрацию мышечных волокон. В результате теплопродукция возрастает на 20-45 % от исходного уровня. При более значительном охлаждении терморегуляционный тонус переходит в мышечную холодовую дрожь. Холодовая мышечная дрожь представляет собой непроизвольную ритмическую активность поверхностно расположенных мышц. В результате теплопродукция возрастает в 2-3 раза по сравнению с исходным уровнем.
Механизмы мышечной дрожи связаны с распространением возбуждения из гипоталамуса через покрышку среднего мозга и через красное ядро к α-мотонейронам спинного мозга и от них - к соответствующим мышцам.
Одновременно при охлаждении в скелетных мышцах, печени и буром жире активируются процессы окисления и снижается эффективность окислительного фосфорилирования. За счет этих процессов, так называемого не сократительного термогенеза, теплопродукция может возрасти в 3 раза.
Регуляция несократительного термогенеза осуществляется активацией симпатической нервной системы, гормонами щитовидной железы и мозгового слоя надпочечников.
№ 55 Механизмы теплоотдачи. Способы отдачи тепла организмом. Физиологические механизмы теплоотдачи.
Поддержание температуры тела на оптимальном для метаболизма уровне осуществляется за счет регулирующего влияния ЦНС. За счет нервных и прямых гуморальных влияний, в которых участвует ряд олигопептидов, например бомбезин, в рассматриваемой функциональной системе формируются процессы, направленные на восстановление сформировавшихся изменений температурной схемы тела. Эти процессы включают механизмы теплопродукции и теплоотдачи.
Центры теплоотдачи. В области передних ядер гипоталамуса обнаружены центры теплоотдачи. Разрушение этих структур приводит к тому, что животные утрачивают способность поддерживать постоянство температуры тела в условиях высокой температуры окружающей среды. Температура их тела при этом начинает возрастать, животные переходят в состояние гипертермии, причем гипертермия может развиться даже при комнатной температуре. Раздражение этих структур через вживленные электроды электрическим током вызывает у животных характерный синдром: одышку, расширение поверхностных сосудов кожи, падение температуры тела. Вызванная предварительным охлаждением мышечная дрожь у них прекращается.
Теплоотдачу (физическую терморегуляцию) определяют физические процессы:
Перемещение теплого воздуха с поверхности тела путем контактной или дистантной конвекции;
Теплоизлучение (радиация);
Испарение жидкости с поверхности кожи и верхних дыхательных
Выделение мочи и кала.
Физическая терморегуляция осуществляется следующими путями.
Контактная конвекция - прямой обмен тепла между двумя объектами с разной температурой, находящимися в прямом контакте друг с другом.
Дистантная конвекция - переход тепла в поток воздуха, который движется около поверхности тела и, нагреваясь, заменяется новым, более холодным.
Радиация - отдача тепла путем излучения электромагнитной энергии в
виде инфракрасных лучей.
Регуляция теплоотдачи. Конвекция, теплоизлучение и испарение тепла прямо пропорциональны теплоемкости окружающей среды.
Теплоотдача зависит от объема поверхности тела. Известно, что многие животные на холоде сворачиваются в клубок, занимая меньший объем. Процессы конвекции, излучения и испарения тепла зависят от свойств кожного покрова. Шерстный покров кожи у животных препятствует теплоотдаче.
Сосудистые реакции при перегревании. В основе всех физических процессов теплоотдачи у человека лежат физиологические процессы, связанные с изменением под влиянием окружающей температуры просвета поверхностных сосудов кожи. При действии высокой температуры сосуды расширяются, при действии низкой - суживаются. Эти реакции осуществляются за счет активации вегетативной нервной системы - парасимпатического отдела в первом случае и симпатического - во втором.
В механизмах расширения сосудов кожи принимает участие брадикинин, который продуцируется потовыми железами через холинергические симпатические волокна.
Теплоотдача в водной среде. Процессы теплоотдачи зависят от физических свойств окружающей среды. Наиболее сложно меняются процессы теплоотдачи, так же как и теплопродукции, в водной среде. Прохладная вода обладает наибольшей теплоемкостью. В воде исключается испарение. Одновременно вода оказывает физическое давление на покровы тела, происходит перераспределение массы тела. Температура воды оказывает раздражающее действие на рецепторы кожи и интерорецепторы.
Потоотделение. Наиболее существенным механизмом теплоотдачи является потоотделение. С 1 г пара организм теряет около 600 кал тепла. Потоотделение имеет существенное значение для поддержания оптимального уровня температуры тела в условиях повышенной температуры окружающей среды, особенно в жарких странах. Установлено, что не все люди в равной степени обладают способностью к усиленному потоотделению в условиях повышенной температуры.
№ 56 Функциональная система, поддерживающая оптимальную для метаболизма температуру крови. Характеристика её узловых механизмов.
Функциональная система, определяющая оптимальную для метаболизма температуру тела, объединяет две подсистемы: внутренней эндогенной саморегуляции и целенаправленного поведения. Эндогенные механизмы саморегуляции за счет процессов теплопродукции и тепловыделения определяют поддержание необходимой для метаболизма температуры тела. Функциональная система:
Полезный приспособительный результат
Показатель, ради которого работает данная функциональная система,- температура крови. С одной стороны, она обеспечивает нормальное течение процессов метаболизма, а с другой - сама определяется их интенсивностью.
Для нормального течения метаболических процессов гомойотермные животные, в том числе и человек, вынуждены поддерживать температуру тела на относительно постоянном уровне. Однако это постоянство условно. Температура различных органов подвержена колебаниям, границы которых зависят от времени суток, функционального состояния организма, теплоизоляционных свойств одежды и др.
Для определения величины энерготрат необходимо знать: (а) какое количество кислорода было потреблено за единицу времени и (б) какое количество энергии должно было образоваться в организме при использовании в окислении1л О 2 - калорический, или энергетический, эквивалент кислорода (КЭК).
Величина КЭК непостоянна. Использование в окислении одного и того же количества кислорода (1 л) приводит к образованию разного количества энергии: чуть больше 5 ккал, если субстратом окисления служат только углеводы;
4,6-4,7 ккал, если окисляются жиры; и 4,5-4,6 ккал при окислении белков.
Поскольку окисление всех трех названных компонентов происходит одновременно, необходимо знать, в каком соотношении они окисляются в данный момент. Для этого определяют дыхательный коэффициент (ДК) - соотношение объемов выделенного углекислого газа и поглощенного за то же время кислорода: ДК = Vco 2 / Vo 2 .
Если бы окислялись исключительно углеводы, то количество молекул кислорода, поглощенных за единицу времени, было бы равно количеству образовавшихся молекул углекислого газа, например: C 6 H 12 O 6 + 60 2 → 6СО 2 + 6Н 2 О.
Равные количества молекул газа занимают равные объемы, следовательно, при окислении углеводов Vco 2 = Vo 2 и ДК =1,0.
В процессе окисления белков часть поглощенного кислорода не выходит затем с выдыхаемым воздухом, но образует соединения с азотом и выводится с мочой. В этом случае ДК принимает значения ниже единицы (примерно 0,81).
При окислении жиров (например, трипальмитина):
2 C 51 H 98 0 6 + 1450 2 → 102С0 2 + 98Н 2 0ДК= 102/145 = 0,70.
Тем не менее, если в течение нескольких недель кормить человека исключительно жирами, ДК не снизится до 0,70. Точно так же, если человек будет питаться чистой глюкозой, ДК (в покое) поднимется только до 0,87, но не до 1,00. Причина - в том, что ежеминутно в окислении используются и углеводы, и жиры, и белки. Компоненты, отсутствующие в пище, мобилизуются из собственных запасов организма. Поэтому ДК обычно принимает некое среднее значение (примерно 0,82).
ДК показывает, в каком соотношении используются в окислении углеводы, жиры и белки в данную минуту. Поэтому к ДК «жестко привязан» КЭК, т. е. каждому значению ДК соответствует своя величина КЭК (табл. 11.1)
Таблица. 11.1
Дыхательный коэффициент (ДК) и калорический эквивалент кислорода (КЭК, ккал/л О 2)
|
дк |
0,70 |
0,75 |
0,80 |
0,85 |
0,90 |
0,95 |
1,00 |
|
КЭК |
4,69 |
4,74 |
4,80 |
4,86 |
4,92 |
4,98 |
5,05 |
При смешанном питании КЭК равен примерно 4,83 ккал/л O 2 . Этой величиной пользуются при приблизительных расчетах (что позволяет не определять экспериментально ДК).
При выполнении человеком тяжелой физической работы накапливающаяся в мышцах молочная кислота вытесняет из крови повышенные количества двуокиси углерода, в результате чего выделение СО 2 нарастает быстрее, чем поглощение О 2 . ДК постепенно приближается к 1, а затем и превышает это значение. ДК =1,12 говорит о значительном ацидозе и о том, что вскоре человек будет вынужден прекратить работу.
После прекращения работы происходит постепенное окисление молочной кислоты, поэтому часть образующегося углекислого газа остается в организме в виде гидрокарбонат-ионов. В результате выделение СО 2 падает быстрее, чем потребление О 2 , и ДК снижается.
Лабораторная работа № 12.
Определение параметров дыхания
Цель работы: познакомиться с основами экспериментального изучения интенсивности дыхания у растений.
Опыт 1. Определение дыхательного коэффициента прорастающих семян
Дыхательный коэффициент (ДК) – это показатель газообмена живых тканей. Он означает отношение количества выделенного при дыхании углекислого газа к количеству поглощенного при этом кислорода:
ДК = СО 2 /О 2 .
Величина дыхательного коэффициента зависит от ряда причин. Первый фактор – химическая природа окисляемого при дыхании субстрата. Если используются углеводы, то ДК близок к единице:
C 6 H 12 O 6 + 6O 2 = 6 CO 2 + 6 H 2 O.
Если окисляются более восстановленные вещества, жиры и белки, то кислорода потребляется больше, чем выделяется углекислого газа, и ДК меньше единицы. Например, при окислении стеариновой кислоты реакция идет по формуле:
C 18 H 36 O 2 +26 O 2 =18 CO 2 +18 CO 2 +18 H 2 0;
и отношение CO 2:O 2 равно 18:26, то есть 0,69.
При окислении веществ, содержащих в себе больше кислорода, чем в углеводах, дыхательный коэффициент больше единицы. Так, при дыхании за счет щавелевой кислоты по уравнению
2C 2 O 2 H 2 + O 2 = 4 CO 2 + 2H 2 O
дыхательный коэффициент равен четырем.
Вторым фактором, определяющим величину ДК, являются условия аэрации. При недостатке кислорода в воздухе, то есть в анаэробных условиях, ДК повышается и в случае окисления углеводов становится выше единицы.
Наконец, величина дыхательного коэффициента свидетельствует о полноте окисления субстрата. Если при окислении углеводов процесс распада идет не до конца, а накапливаются промежуточные, более окисленные, чем углеводы, продукты, то величина ДК становится меньше единицы. Подобное явление наблюдается у интенсивно растущих объектов. В работе предлагается один из наиболее простых методов определения ДК прорастающих семян – метод Рихтера.
Ход работы: в опыте используют прибор, состоящий из пробирки, которая плотно закрыта каучуковой пробкой, со вставленной в неё горизонтальной трубкой с делениями. Пробирку поместить в колбу, которая является одновременно и штативом, и термоизолятором.
Прорастающими семенами пшеницы или подсолнечника заполнить 1/2–2/3 объема пробирки и плотно закрыть ее пробкой с измерительной трубкой. Обязательное условие правильного наблюдения – постоянство температуры прибора, так как его работа связана с изменением объемов газов.
Поэтому смонтированный прибор должен принять комнатную температуру, что достигается в течение 5–7 минут.
В конец измерительной трубки ввести каплю жидкости (например, подкрашенную метиленовой синью воду). Для этого, не вынимая пробирку из колбы, погрузить конец трубки в стаканчик с жидкостью. Если жидкость плохо поступает в отверстие, можно слегка постучать кончиком трубки о дно стакана или, вынув трубку из стакана, протереть кончик её фильтровальной бумагой. Если эти меры не помогают, то надо капилляр трубки промыть спиртом и водой. Капля должна подняться по трубке на расстояние 1 см. Таким образом, в приборе создается замкнутое пространство. Всякое изменение в нем объема газов приведет каплю в движение. По скорости этого движения можно судить о газообмене.
При равенстве объемов выделяющегося углекислого газа и поглощающегося кислорода общий объем газов в пробирке останется неизменным, и капля не будет менять своего положения. В том случае, когда поглощается больше О 2 , чем выделяется СО 2 , в пробирке возникает разряжение газов и капля передвигается внутри по трубке. При условии более интенсивного выделения СО 2 по сравнению с поглощением кислорода капля будет выбрасываться из трубки. Требуется пронаблюдать за поведением капли и, если она перемещается, рассчитать скорость ее движения, определяемую разностью объемов О 2 и СО 2 . С этой целью, как только в кончик трубки будет введена капля, необходимо засечь время и ждать, через сколько минут капля пройдет какой-то отрезок пути S . Зная путь S и время t , затраченное на преодоление этого пути, можно рассчитать скорость движения капли:
V = S/ t .
Эту скорость обозначить буквой А. Определить ее 2–3 раза, для дальнейших расчетов взять среднюю величину.
Затем пробирку раскрыть и ввести в нее фильтровальную бумажку, смоченную концентрированным раствором щелочи, которая будет поглощать выделяющийся при дыхании углекислый газ. Пробирку снова закрыть пробкой, дать прибору принять комнатную температуру и 2–3 раза определить скорость движения капли после введения щелочи, обозначить её В. Она зависит от объема поглощенного при дыхании семян кислорода: В = О 2 .
Таблица 1
Схема записи опыта
Задание: Значения А и В записать в таблицу 1 и рассчитать величину дыхательного коэффициента объекта, используемого в опыте. Расчет делать по формуле: ДК = В / (В− А). Эта дробь дает величину отношения СО 2 к О 2 , т. к. В = О 2 ; А = О 2 – СО 2 .
В конце работы объяснить, от чего зависит полученная величина ДК.
Опыт 2. Упрощенный метод определения интенсивности дыхания (по Бойсен-Иенсену)
Настоящий метод основан на учете количества углекислого газа, выделяемого растением в замкнутом пространстве и поглощаемого баритом.
Удобным объектом для работы служат семена. Нужно сравнить интенсивность дыхания прорастающих и покоящихся семян пшеницы.
Следует иметь в виду, что у этого метода есть недостаток: объект находится в замкнутой атмосфере, где может возникнуть дефицит кислорода, и в парах барита, ядовитого для организмов.
Материал и объекты исследования: конические колбы емкостью на 100–200 мл, пробки резиновые, марля для мешочков, нитки, бюретки для титрования, штативы, весы, 0,1 N раствор Ва(ОН) 2 , 0,1 N раствор щавелевой кислоты, фенолфталеин, проросшие и покоящиеся семена пшеницы, гороха, ячменя, подсолнечника и др.
Ход работы: в три конические колбы с пробками емкостью на 100–200 мл с помощью бюретки налить по 10 мл 0,1N Ва(ОН) 2 . Барит ядовит для человека, и пипеткой брать его небезопасно! Колбы закрыть пробками, чтобы избежать попадания в барит углекислоты из воздуха.
На технических весах взять навески по 3 г сухих и прорастающих семян и поместить их в марлевые мешочки. Мешочки подвесить в колбы над баритом с помощью крючков, вставленных в резиновые пробки.
Необходимо следить, чтобы марля не касалась раствора барита. Третью колбу (без семян) использовать в качестве контроля.
Экспозиция опыта не должна превышать одного часа, так как семена могут оказаться в анаэробных условиях. В ходе опыта колбы надо аккуратно покачивать, чтобы разрушить пленку углекислого бария, образующегося на поверхности раствора при поглощении СО 2 . Время опыта должно быть точно учтено.
В конце опыта барит в колбах оттитровать 0,1 N раствором щавелевой кислоты. Титрование лучше начать с контрольной колбы. Сначала установить мениск раствора щавелевой кислоты в бюретке, только после этого открыть пробку и вынуть мешочек с семенами. В раствор барита внести 2–3 капли фенолфталеина, который в щелочи приобретает розовую окраску. Титрование проводить по возможности быстро, чтобы барит не успел поглотить много СО 2 из воздуха. Конец титрования определяется по обесцвечиванию фенолфталеина от одной капли щавелевой кислоты. Раствор становится молочно-белым, и при дальнейшем прибавлении щавелевой кислоты окраска его не меняется. Поэтому важно не пропустить конец титрования. Для проверки точности титрования в обесцвеченный раствор можно капнуть бариту: при хорошей работе от одной капли Ва(ОН) 2 раствор снова порозовеет.
Расчет интенсивности дыхания сделать по формуле
J = / (nt ), мг СО 2 /г/ч,
где а – количество щавелевой кислоты, пошедшей на титрование при контроле, мл; b – количество щавелевой кислоты, пошедшей на титрование в опытном варианте, мл; 2,2 – мг СО 2 , соответствует 1 мл 0,1 N щавелевой кислоты; n – вес сухих семян, г; t – время опыта, мин.
Результаты титрования и расчетов записать в таблицу 2.
Таблица 2
Схема записи опыта
Задание: в конце работы сделать вывод о зависимости интенсивности дыхания от влажности семян.
Читатель, наверное, заметил, что в норме каждые 100 мл крови переносят от легких
в ткани около 5 мл кислорода, в то же время из тканей в легкие переносится только около 4 мл двуокиси углерода. Таким образом, в нормальных условиях в покое объем выведенного через легкие двуокиси углерода составляет только 82% объема поглощенного в легких кислорода. Соотношение количества выведенной двуокиси углерода к количеству поглощенного кислорода называют дыхательным коэффициентом (ДК).
Дыхательный коэффициент
= Объем выведенной двуокиси углерода / Объем поглощенного кислорода.
При изменении условий обмена величина дыхательного коэффициента изменяется. При использовании в пище только углеводов величина ДК достигает 1, и наоборот, при использовании в пище только жиров величина ДК снижается до 0,7. Причина этих изменений заключается в том, что при использовании одной молекулы кислорода в метаболизме углеводов образуется одна молекула двуокиси углерода; при реакции кислорода с жирами большое количество кислорода соединяется с атомами водорода жира, и вместо двуокиси углерода образуется вода. Другими словами, при обмене жиров дыхательный коэффициент химических реакций в тканях составляет около 0,7 вместо 1. Считается, что для человека, питающегося пищей, содержащей средние количества углеводов, жиров и белков, средняя величина ДК составляет 0,825.
Дыхательный центр
Нервная система обычно устанавливает такую скорость альвеолярной вентиляции, которая почти точно соответствует потребностям тела, поэтому напряжение кислорода (Ро2) и двуокиси углерода (Рсо2) в артериальной крови мало изменяется даже при тяжелой физической нагрузке и при большинстве других случаев респираторного стресса. В этой главе изложена функция этой нейрогенной системы регуляции дыхания.
Дыхательный центр состоит из нескольких групп нейронов, расположенных в стволе мозга по обе стороны продолговатого мозга и моста. Их делят на три большие группы нейронов: (1) дорсальная группа дыхательных нейронов, расположенная в дорсальной части продолговатого мозга, которая в основном вызывает вдох; (2) вентральная группа дыхательных нейронов, которая расположена в вентролатеральной части продолговатого мозга и в основном вызывает выдох; (3) пневмотаксический центр, который расположен дорсально в верхней части моста и контролирует в основном скорость и глубину дыхания. Наиболее важную роль в контроле дыхания выполняет дорсальная группа нейронов, поэтому первой будем рассматривать ее функции.
Дорсальная группа дыхательных нейронов простирается на большую часть длины продолговатого мозга. Большинство этих нейронов расположено в ядре одиночного тракта, хотя расположенные в близлежащей ретикулярной формации продолговатого мозга дополнительные нейроны также имеют важное значение для регуляции дыхания. Ядро одиночгого тракта является сенсорным ядром для блуждающего и языкоглоточного нервов, которые передают в дыхательный центр сенсорные сигналы от: (1) периферических хеморецепторов; (2) барорецепторов; (3) разного типа рецепторов легких.
Ритмические инспираторные разряды от дорсальной группы нейронов. Базовый ритм дыхания генерируется в основном дорсальной группой дыхательных нейронов. Даже после перерезки всех входящих в продолговатый мозг периферических нервов и ствола мозга ниже и выше продолговатого мозга эта группа нейронов продолжает генерировать повторяющиеся залпы потенциалов действия инспираторных нейронов. Основная причина возникновения этих залпов неизвестна. У примитивных животных были найдены нервные сети, в которых активность одной группы нейронов активирует вторую группу, а активность второй группы тормозит первую.
Через некоторое время схема активации повторяется , и так продолжается в течение всей жизни животного, поэтому большинство физиологов, занимающихся физиологией дыхания, полагают, что у человека тоже имеется подобная сеть нейронов, расположенная в пределах продолговатого мозга; возможно, что в нее входит не только дорсальная группа нейронов, но и прилегающие части продолговатого мозга, и что эта сеть нейронов отвечает за основной ритм дыхания.
Нарастающий сигнал вдоха
. Сигнал от нейронов, который передается инспираторным мышцам, в основном диафрагме, не является мгновенным всплеском потенциалов действия. При нормальном дыхании он постепенно увеличивается в течение примерно 2 сек. После этого он резко снижается примерно на 3 сек, что прекращает возбуждение диафрагмы и позволяет эластической тяге легких и грудной стенки выполнить выдох. Потом инспираторный сигнал начинается опять, и цикл повторяется снова, и в перерыве между ними происходит выдох. Таким образом, инспираторный сигнал является нарастающим сигналом. По-видимому, такое нарастание сигнала обеспечивает постепенное увеличение объема легких во время вдоха вместо резкой инспирации.
Контролируются два момента
нарастающего сигнала.
1. Скорость прироста нарастающего сигнала, поэтому во время затрудненного дыхания сигнал растет быстро и вызывает быстрое наполнение легких.
2. Лимитирующая точка, при достижении которой сигнал внезапно пропадает. Это обычный способ контроля над скоростью дыхания; чем раньше прекращается нарастающий сигнал, тем меньше длительность вдоха. При этом сокращается и длительность выдоха, в результате дыхание учащается.
Азотистый баланс - соотношение количества азота, поступившего в организм с пищей и выделенного из него. Так как основным источником азота в организме является белок, то по азотистому балансу можно судить о соотношении количества поступившего и разрушенного в организме белка. Количество принятого с пищей азота отличается от количества усвоенного азота, так как часть азота теряется с калом.
Усвоение азота вычисляют по разности содержания азота в принятой пище и в кале. Зная количество усвоенного азота, легко вычислить общее количество усвоенного организмом белка, так как в белке содержится в среднем 16% азота, т. е. 1 г азота содержится в 6,25 г белка. Следовательно, умножив найденное количество азота на 6,25, можно определить количество белка.
Для того чтобы установить количество разрушенного белка, необходимо знать общее количество азота, выведенного из организма. Азотсодержащие продукты белкового обмена (мочевина, мочевая кислота, креатинин и др.) выделяются преимущественно с мочой и частично с потом. В условиях обычного, неинтенсивного потоотделения на количество азота в поте можно не обращать внимания. Поэтому для определения количества распавшегося в организме белка обычно находят количество азота в моче и умножают на 6,25.
Между количеством азота, введенного с белками пищи, и количеством азота, выводимым из организма, существует определенная взаимосвязь. Увеличение поступления белка в организм приводит к увеличению выделения азота из организма. У взрослого человека при адекватном питании, как правило, количество введенного в организм азота равно количеству азота, выведенного из организма. Это состояние получило название азотистого равновесия. Если в условиях азотистого равновесия повысить количество белка в пище, то азотистое равновесие вскоре восстанавливается, но уже на новом, более высоком уровне. Таким образом, азотистое равновесие может устанавливаться при значительных колебаниях содержания белка в пище.
В случаях, когда поступление азота превышает его выделение, говорят о положительном азотистом балансе. При этом синтез белка преобладает над его распадом. Устойчивый положительный азотистый баланс наблюдается всегда при увеличении" массы тела. Он отмечается в период роста организма, во время беременности, в периоде выздоровления после тяжелых заболеваний, а также при усиленных спортивных тренировках, сопровождающихся увеличением массы мышц. В этих условиях происходит задержка азота в организме (ретенция азота}.
Белки в организме не депонируются, т. е. не откладываются в запас. Поэтому при поступлении с пищей значительного количества белка только часть его расходуется на пластические цели, большая же часть - на энергетические цели.
Когда количество выведенного из организма азота превышает количество поступившего азота, говорят об отрицательном азотистом балансе.
Отрицательный азотистый баланс отмечается при белковом голодании, а также в случаях, когда в организм не поступают отдельные необходимые для синтеза белков аминокислоты.
Распад белка в организме протекает непрерывно. Степень распада белка связана с характером питания. Минимальные затраты белка в условиях белкового голодания наблюдаются при питании углеводами. В этих условиях выделение азота может быть в 3-3"/2 раза меньше, чем при полном голодании. Углеводы при этом выполняют сберегающую белки роль.
Распад белков в организме, происходящий при отсутствии белков в пище и достаточном введении всех других питательных веществ (углеводы, жиры, минеральные соли, вода, витамины), отражает те минимальные траты, которые связаны с основными процессами жизнедеятельности. Эти наименьшие потери белка для организма в состоянии покоя, пересчитанные на 1 кг массы тела, были названы Рубнером коэффициентом изнашивания.
Коэффициент изнашивания для взрослого человека равен 0,028-0,075 г азота на 1 кг массы тела в сутки.
Отрицательный азотистый баланс развивается при полном отсутствии или недостаточном количестве белка в пище, а также при потреблении пищи, содержащей неполноценные белки. Не исключена возможность дефицита белка при нормальном поступлении, но при значительном увеличении потребности в нем организма. Во всех этих случаях имеет место белковое голодание. "
При белковом голодании даже в случаях достаточного поступления в организм жиров, углеводов, минеральных солей, воды и витаминов происходит постепенно нарастающая потеря массы тела, зависящая от того, что затраты тканевых белков (минимальные в этих условиях и равные коэффициенту изнашивания) не компенсируются поступлением белков с пищей. Поэтому длительное белковое голодание в конечном счете, так же как и полное голодание, неизбежно приводит к смерти. Особенно тяжело переносят белковое голодание растущие организмы, у которых в этом случае происходит не только потеря массы тела, но и остановка роста, обусловленная недостатком пластического материала, необходимого для построения клеточных структур.